Geographic Distribution and Genetic Diversity of Podosphaera xanthii in Pakistan
-
Najam-ul-Sehar Afshan
Institute of Botany, University of the Punjab, Lahore 54590, Punjab, Pakistan
Muhammada Jabeen
Institute of Botany, University of the Punjab, Lahore 54590, Punjab, Pakistan
Afshan AkbarDepartment of Botany, Hazara University, Mansehra 21300, Khyber Pakhtunkhwa, Pakistan
Muhammadah KhalidInstitute of Botany, University of the Punjab, Lahore 54590, Punjab, Pakistan
Arooma SaleemInstitute of Botany, University of the Punjab, Lahore 54590, Punjab, Pakistan
Rabia AltafInstitute of Botany, University of the Punjab, Lahore 54590, Punjab, Pakistan
Saliha AfzalInstitute of Botany, University of the Punjab, Lahore 54590, Punjab, Pakistan
Muhammad FiazDepartment of Botany, Hazara University, Mansehra 21300, Khyber Pakhtunkhwa, Pakistan
Abdul Nasir KhalidInstitute of Botany, University of the Punjab, Lahore 54590, Punjab, Pakistan
| Received 11 Apr, 2025 |
Accepted 01 Aug, 2025 |
Published 10 Aug, 2025 |
Background and Objective: Podosphaera xanthii is one of the most commonly occurring powdery mildew pathogens, with a broad host range and wide distribution. Despite its prevalence, an updated record of its host plants in Pakistan was lacking. This study aimed to update the host range of P. xanthii based on recent surveys, confirm its occurrence through morphological and molecular analyses, and highlight its diversity and regional spread in Pakistan. Materials and Methods: Field surveys were conducted across five regions of Pakistan Battagram, Fairy Meadows, Khanspur, Mansehra, and Swat between 2022 and 2024. Infected plant samples were collected and analyzed morphologically, focusing on both anamorphic and teleomorphic stages. Molecular confirmation was done through sequencing of the internal transcribed spacer (ITS) region. Descriptive statistics were used to report host range diversity and geographic occurrence. Results: A total of ten plant species were confirmed as hosts of P. xanthii, based on morphological characteristics and ITS-based molecular validation. Among these, members of the Asteraceae family were dominant. The fungus was found in all surveyed regions, indicating a wide ecological range and adaptability. The simultaneous observation of both reproductive stages supports its active and persistent lifecycle across diverse climatic zones. Conclusion: This study provides an updated account of P. xanthii host plants in Pakistan, confirming its widespread occurrence and significant diversity. The findings underscore the need for targeted management strategies to mitigate its impact, especially on susceptible plant families like Asteraceae.
INTRODUCTION
Powdery mildew fungi belonging to the family Erysiphaceae are obligate biotrophic parasites that grow on both sides of leaf surfaces, petioles, and stems. They are characterized by a whitish powdery mass that obtains nutrients by means of haustoria1. They may develop in open fields and greenhouses and reduce the crop yields up to 40%2. The family Erysiphaceae is composed of five major tribes, including Erysipheae Tul. & C. Tul., Golovinomyceteae U. Braun, Cystotheceae Katumoto, Phyllactinieae Palla, and Blumerieae Cook. The tribe Cystotheceae is further composed of Cystotheca Berk. and Curtis, Podosphaera Kunze, and Sawadaea Miyabe. Podosphaera is a pathogenic fungus of herbaceous plants and trees, while Cystotheca, Sawadaea are pathogens of trees3.
The genus Podosphaera is capable of both sexual and asexual reproduction. In the teleomorphic stage, fruiting bodies (chasmothecia) develop and contain a single ascus. Formation of a single ascus is the distinguishing feature of the genus Podosphaera, which distinguishes it from other powdery mildew fungi. Another distinguishing teleomorphic character of Podosphaera is the formation of myceloid, uncinate, circinate, or dichotomously branched hyphal appendages. Anamorphic characters of Podosphaera include the formation of multiple conidiospores per condiophore and are almost similar to other genera of Erysiphaceae4-7.
Among various Podosphaera species, P. xanthii mostly infects trees and herbaceous plants and is reported on numerous hosts of the family Asteraceae, Balsaminaceae, Fabaceae, Solanaceae, Verbenaceae in North, South America, Asia, Australia, Africa, Europe, and New Zealand3. This fungus is causing enormous economic losses in members of the family Cucurbitaceae. The nomenclature and host range of P. xanthii have changed repeatedly and were reported for the first time under the name Sphaerotheca fuliginea Schltdl. by Sawada in 1919, causing infection on 15 different plant species. Braun in 1987 reduced this species to Sphaerotheca fusca (Fr.) S. Blumer and in 2001 re-introduced this species as P. xanthii after examining its infection on hosts of various plant families3-8. Despite the widespread occurrence of Podosphaera xanthii and its economic impact, data on its host range and distribution in Pakistan remain sparse. In this study, new host records of P. xanthii from Pakistan based on recent field collections have been documented. These records are supported by morphological characterization of both anamorphic and teleomorphic stages, as well as molecular identification using DNA sequence analysis. This work contributes to the understanding of the biodiversity, host specificity, and biogeography of P. xanthii, and serves as a foundation for future research on its management and taxonomy.
MATERIALS AND METHODS
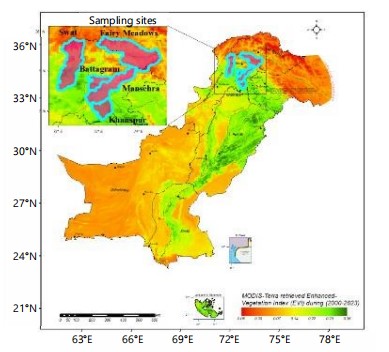
Collection and preservation: In the years 2022-2024, the symptoms of powdery mildew disease were observed on the leaves of Bidens alata Melchert., Chrysanthemum coronarium L., Cirsium arvense (L.) All, Cucurbita maxima L., C. pepo L., Impatiens bicolor Royle., Momordica charantia L., Salvia officinale (L.) All. Senecio chrysanthimoides DC., and Xanthium strumarium L. during phytopathological surveys conducted in Battagram, Fairy Meadows, Khanspur, Mansehra, Swat, Pakistan. Infected plant samples were collected and shade-dried on blotting papers. Then, these were preserved in paper envelopes and deposited in the herbarium of the Institute of Botany, University of the Punjab, Lahore (LAH38549, LAH38326, LAH38560, LAH38202, LAH38037, LAH39550) and Department of Botany, Hazara University, Mansehra (HUP16751, HUP16733, HUP16768, HUP16753). Sampling sites are featured in Fig. 1.
Microscopic analysis: Host plants were identified by comparison with healthy preserved plants present in the Herbarium, Hazara University, Mansehra (HUP); Institute of Botany, University of the Punjab, Lahore (LAH). Morphological features were observed under a stereomicroscope (EMZ-5TR, Meiji Techno Co., Ltd., Japan). Conidiophores, conidia, foot cells, and appressoria were stripped from the leaf surface with a clean needle and mounted in lactic acid. The slides were observed under a light microscope (LABOMED, Labo America, Inc., USA) with attached camera (HDCE-90D). Measurements of microscopic features were taken with the help of Scope Image 9.0(X5). Twenty-five measurements each for anamorph (conidiophores, conidia, foot cells, appressoria) and teleomorph (chasmothecia, asci, ascospores) were made after observing their shapes.
Molecular analysis: Fungal mycelia were scratched from the infected plant samples while observing under the stereomicroscope. Autoclaved and ice-chilled pestles and mortars were used along with liquid nitrogen for crushing the material and were preserved in Eppendorf tubes at -18°C.
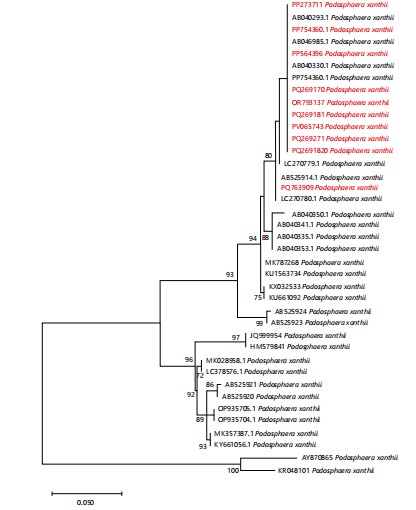
Genomic DNA was extracted using GeneJET Plant Genomic DNA Purification Mini Kit. #K0791 (Thermo Scientific, Vilnius, Lithuania) according to the manufacturer’s instructions. The Internal Transcribed Spacer (ITS) region was amplified using primers i.e, Forward primer PMITS1 (5´-TCGGACTGGCC(T/C)AGGGAGA-3´) and Reverse primer PMITS2 (5´-TCACTCGCCGTTACTGAGGT-3´)9. Sanger dideoxy sequencing was done by Tsingke in China. Consensus was made using forward and reverse sequences on Bioedit10. Consensus sequences were then run in NCBI BLAST to search and collect similar sequences of the ITS regions from the GenBank database. Sequences of different published Podosphaera taxa were obtained from GenBank to determine the phylogenetic position of the sample. Multiple sequence alignment was performed using MAFFT. Trimming of sequences was done using BioEdit at their conserved sites. Software MEGA 7.0 was used for the reconstruction of a phylogenetic tree based on the Kimura 2-parameter model11. One thousand bootstrap replicates were run to infer the evolutionary history of the species. Phyllactinia guttata and Phyllactinia populi were used as an outgroup (Fig. 2).
|
RESULTS AND DISCUSSION
Podosphaera xanthii (Castagne) U. Braun & Shishkoff, Schlechtendalia 4: 31, 2000: First record of Bidens alata melchert (Asteraceae) in Pakistan (Fig. 3a-i).
Material examined: On Bidens alata Melchert. (Asteraceae), with anamorphic stage, Battagram, Khyber Pakhtunkhwa, Pakistan, 1850 m.a.s.l., 13th October, 2022, AFE072, Voucher No. HUP 16751, GenBank Accession No. PQ2691820 (ITS).
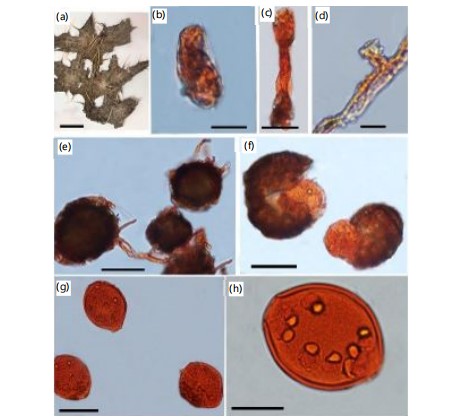
Mycelium amphigenous, stems, in irregular patches or effuse, thin to dense, evanescent to±persistent; hyphae thin walled, smooth or almost so, (3-)5-8(-10) μm wide; chasmothecia scattered to gregarious, (70-)80-110(-115) μm diam (Fig. 3c-e); peridium cells conspicuous, large, (10-)15-55 μm diam., shape irregularly polygonal to daedaleoid, i.e. with sinuous walls; chasmothecial appendages few to numerous, in the lower half, mycelioid, simple or irregularly branched, often interwoven with each other and the mycelium, length variable, 4-12 μm wide, hyaline, later brown throughout or paler towards the tips, short, coarse, brown appendages often mixed with longer, paler appendages, septate, walls thin, smooth to rough (Fig. 3d); ASCUS broadly ellipsoid-ovoid to subglobose, 60-80×50-65 μm, sessile, wall up to 3 μm thick, terminal oculus (-10)15-25 μm diam., (6-)8-spored, mostly immature (Fig. 3f); ascospores broadly ellipsoid-ovoid to subglobose, 15-23×12-17 μm, maturing late, but occasionally larger when fully mature, colourless (Fig. 3f); conidiophores arising from the upper surface of hyphal mother cells, mostly towards one end of the cell but sometimes±centrally, erect, foot cells cylindrical, 30-100×10-13 μm, sometimes slightly constricted at the basal septum or slightly swollen at the very base, followed by 1-3(-4) shorter cells, forming catenescent conidia, in long chains (Fig. 3g); conidia ellipsoid-ovoid to doliiform, 25-45×14-22 μm (Fig 3h-i).
|

|
The ITS region showed 99.38% similarity with P. xanthii reported from Japan12. Previously, P. xanthii was recorded on Bidens L. plant in Korea, Thailand and Taiwan8,13,14. Bidens plant was also infected by other powdery mildew species, including P. bidenticola U. Braun on the morphological basis3. Bidens alata is a new host record for P. xanthii from Pakistan.
First record on Cirsium arvense (L.) All. (Asteraceae) in Pakistan (Fig. 4a-h).
Material examined: On Cirsium arvense (L.) All. (Asteraceae) with Anamorph & Teleomorph, Mansehra, Hazara, Khyber Pakhtunkhwa, Pakistan, 3140 m.a.s.l., 21 August, 2023, MK 100, Voucher No. LAH 38450, Genbank Accession No. PP754360.1 (ITS).
Mycelium amphigenous, on stem and inflorescences, irregular patches, thin to dense; hyphal walls thin walled, smooth, 3-6 μm wide; hyphal appressoria slightly lobed; conidiophores centrally erect, septate, 40-90 μm long, with constricted cylindrical foot cells, with 2-4 following cells, forming catenescent, doliiform, conidia; 10-13 (12)×27-39 (33) μm (Fig. 4c); chasmothecia scattered to gregarious, 75-105 (90) μm diameter (Fig. 4e); peridium cells large, daedaleoid, 20-45 μm in diameter (Fig. 4f); appendages mycelioid, few to abundant, smooth, simple, irregularly branched, dense at base, paler towards tip, short dense appendage interwoven with longer and pale appendages, 4-8 (7.5)×30-50 (42.5) μm; ASCI ellipsoid-ovoid, subglobose, sessile or very short stalked saccate, 7-8 spored, 52-66 (65)×57-79 (77) μm (Fig. 4f).
|
The ITS sequence was up to 100 % identical to published ones AB046985.1 in GenBank15. Cirsium arvense for P. xanthii is a new host record for Pakistan. Cirsium was recorded for P. xanthii in Korea16. Powdery mildew is frequently observed on this host in Pakistan. Cirsium has been reported as a host plant for Erysiphe mayorii S. Blumer, Golovinomyces biocellatus Ehrenb., G. depressus Wallr., G. montagnei U. braun, Leveillula duriaei Lev., L. lappae Castagne, whereas Cirsium species in New Zealand are infected by P. xanthii3.
First record on Cucurbita maxima L. (Asteraceae) in Pakistan (Fig. 5a-k).
Material examined: On leaves of Cucurbita maxima L. (Cucurbataceae), with only anamorphic stage, Fairy Meadows, Diamir, Gilgit Baltistan, Pakistan, 3,300 m.a.s.l., August 06, 2023, FM 12, Voucher No. LAH38326 GenBank accession number PQ763909 (ITS); on leaves of Cucurbita pepo L. (Cucurbataceae), with only anamorphic stage, Khanaspur, KPK, Pakistan, October 10, 2024, RA24, Voucher No. LAH38560 GenBank accession number PV065743 (ITS).
|
Mycelium present on both upper and lower surfaces and on inflorescences and stems, in effuse or irregular patches, dense to thin, persistent to evanescent; hyphae smooth, thin-walled; appressoria slightly lobed shaped to indistinct, solitary (Fig. 5d); conidiophores erect, emerging from the top surface of hyphal mother cells, towards one end of the cell mostly (Fig. 5b); foot cells cylindrical, 30-100×10-13 μm, swollen at the very base, having 1-3(-4) following shorter cells, forming canescent conidia, in long chains (Fig. 5c-h); CONIDIA doliiform to ellipsoid-ovoid, 25-45×14-22 μm (Fig. 5e-g); GERM TUBES are of Fibroidium type.
The ITS sequence was 100% identical to P. xanthii (OQ729760) from Turkey in GenBank. Cucurbita maxima has been reported here as a new host plant for P. xanthii from Pakistan. Previously, Cucurbita was recorded as a host plant for Golovinomyces cucurbitacearum, Leveillula taurica and Podosphaera xanthii on a morphological basis3.
First record on Chrysanthemum coronarium L. (Asteraceae) in Pakistan (Fig. 6a-f).
Material examined: On Chrysanthemum coronarium L. (Asteraceae), with anamorph, Swat, Khyber Pakhtunkhwa, Pakistan, 2409 m.a.s.l, 10 September 2022, DS–19, Voucher No. LAH38202, GenBank accession number PP564396 (ITS).

|

|
Mycelium dense, persistent, amphigenous, forming white patches; chasmothecia scattered, globose to sub-globose, brown, 70-115 μm diam (Fig. 6c); peridial cells irregularly shaped, 15-50 μm diam (Fig. 6c); ascus sessile, globose to ellipsoid, immature, 60-90×60-80 μm, 6-8 spored (Fig. 6e); ascospores hyaline, ellipsoid to ovoid, 15-22×13-18 μm (Fig. 6f).
About 904 species of powdery mildews belonging to 16 genera are reported worldwide17, while from Pakistan, 52 species of powdery mildews are known18-26. The genus Podosphaera Kunze comprises 146 species that cause disease on 1,056 plant species of 41 families22. The combination of ITS-based phylogeny and morpho-anatomical description suggested that the identity of our collection is Podosphaera xanthii (PP564396). Chrysanthemum coronarium is being reported here as a new host plant for P. xanthii from Pakistan and worldwide.
First record on Impatiens bicolor Royle. (Boraginaceae) from Pakistan (Fig. 7a-f).
Material examined: On Impatiens bicolor Royle. (Boraginaceae), with anamorphic stage, Battagram, Hazara, Khyber Pakhtunkhwa, Pakistan, 1430 m.a.s.l., 13th September, 2021, AFE021, Voucher No. HUP 16733, Gen Bank Accession number (PQ269181).
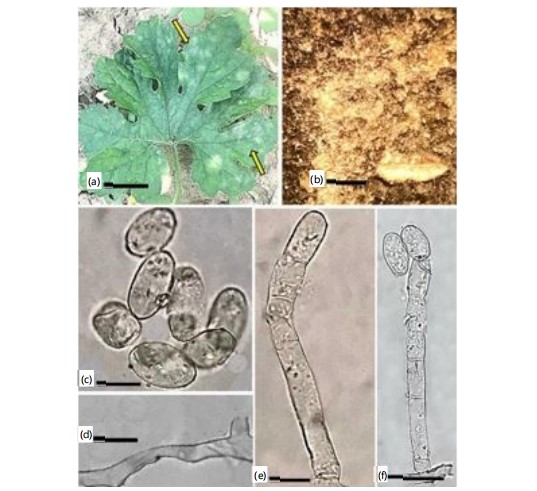
|
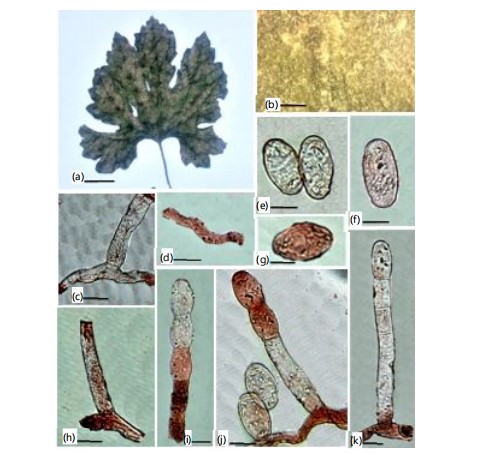
Mycelium amphigenous and on stems and inflorescences, in irregular patches or effuse, thin to dense, evanescent to±persistent; conidiophores arising from the upper surface of hyphal mother cells, mostly towards one end of the cell but sometimes±centrally, erect, footcells cylindrical, 30-100×10-13 μm, sometimes slightly constricted at the basal septum or slightly swollen at the very base, followed by 1-3(-4) shorter cells, forming catenescent conidia, in long chains (Fig. 7c-d); CONIDIA ellipsoid ovoid to doliiform, 25-45×14-22 μm (Fig. 7e-f).
Impatiens plant was recorded as a host plant for a wide range of powdery mildews, including Fibroidium abelmoschi Thum, Podosphaera balsaminae Wallr., P. bidenticola U. Braun, and P. xanthii Castagne. Impatiens bicolor has been reported here as a new host plant for P. xanthii from Pakistan.
First record on Momordica charantia L. (Cucurbitaceae) in Pakistan (Fig. 8a-f).
Material examined: On Momordica charantia L. (Cucurbitaceae), with anamorph, Pakistan, Khyber Pakhtunkhwa, Shogran, 2,362 m.a.s.l., September 29, 2021, SA-01, Voucher No. LAH38037, GenBank accession numbers OR793137 (ITS).
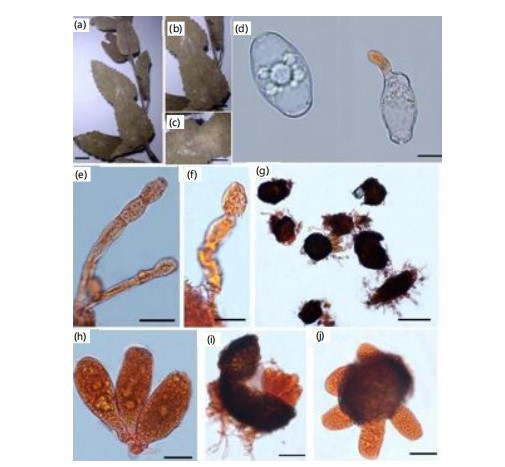
|
Mycelium amphigenous, patchy, thin, forming irregular white, evanescent. Hyphal walls branched, septate, hyaline, thin-walled, smooth, sub-straight to sinuous, 5-8 μm wide. Hyphal appressoria solitary, slightly nipple-shaped, 3-5 μm in diameter (Fig. 8d). Conidiophores hyaline, two to four immature conidia produced in chains with a sinuate outline, erect, straight, 93.7-142.2 (110.2) μm long, followed by one to three cells (Fig. 8e-f). Foot cells are clear, usually straight, sometimes slightly constricted at the basal septum or slightly swollen at the very base, cylindrical or subcylindrical, elongated, 55.2-75.5 (66.3) μm long, forming canescent conidia (Fig. 8e-f). Conidia hyaline, vacuolated, ellipsoid–ovoid, doliiform, smooth, primary conidia were rounded at the apex and subtruncate at the base, 29.0-38.0 (27.0)×16.0-20.0 (29.0) μm (Fig. 8c). Fibrosin bodies present. Chasmothecium was not observed.
Podosphaera xanthii has been infecting Momordica charantia L., a new host record for Pakistan. Previously, the fungus has been reported on Ageratum conyzoides L. and Tarlmounia elliptica (DC.) H. Rob., S.C. Keeley, Skvarla & R. Chan in Lahore18, from the country. Another powdery mildew, Oidium xanthii Bhatn & Kothari, has been found on Xanthium strumarium L. in India. It is morphologically similar to the P. xanthii anamorph, but has conidia with fibrosin bodies, distinguishing it from the former3.
First record on Salvia officinalis (L.) All., (Lamiaceae) in Pakistan (Fig. 9a-j).
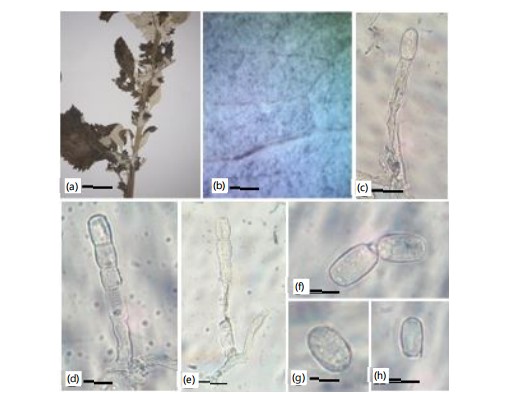
|
Material examined: On Salvia officinale (L.) All. (Lamiaceae), with Anamorph & Teleomorph, Pakistan. Khyber Pakhtunkhwa, Hazara, District Mansehra, Burwai, 3140 m.a.s.l., August, 2023, MK06, Voucher No. LAH38550, GenBank accession number PQ296170 (ITS).
Mycelium amphigenous, patchy, thin, forming irregular white, evanescent. Hyphal walls branched, septate, hyaline, thin-walled, smooth, sub-straight to sinuous, 4-7 μm wide. Hyphal appressoria solitary, slightly nipple-shaped, 3-5 μm in diameter. Conidiophores hyaline, two to four immature conidia produced in chains with a sinuate outline, erect, straight, 94.7-150.2 (114.2) μm long, followed by one to three cells (Fig. 9e-f). Foot cells are clear, usually straight, sometimes slightly constricted at the basal septum or slightly swollen at the very base, cylindrical or subcylindrical, elongated, 57.2-79.5 (70.3) μm long, forming canescent conidia. Conidia hyaline, vacuolated, ellipsoid-ovoid, doliiform, smooth, primary conidia were rounded at the apex and subtruncate at the base, 30.0-39.0 (28.0)×14.0-21.0 (27.0) μm (Fig. 9d).
Podosphaera xanthii has been infecting Salvia officinale, a new host record for Pakistan. Previously, the fungus has been reported on Ageratum conyzoides L. and Tarlmounia elliptica (DC.) H. Rob., S.C. Keeley, Skvarla & R. Chan in Lahore27, from the country. The fungus showed a white mat of mycelium on both sides of the leaf. Black dots were also present on the leaf. Conidia and conidiophores of powdery mildew were verified by microscopic examination. Straight, unbranched conidiophores with cylindrical foot cells that measured 40-80×8-12 μm, followed by one to three smaller cells. They ranged in shape from ellipsoid to ovoid or doliform and were 20-35×14-20 μm3.
First record on Senecio chrysanthimoides (Asteraceae) in Pakistan (Fig. 10a-h).
Material examined: On Senecio chrysanthimoides (Asteraceae) with anamorphic stage, Battagram, Hazara, Khyber Pakhtunkhwa, Pakistan, 1820 m.a.s.l., 15th September, 2022, AFE099, Voucher No. HUP16768, GenBank Accession No. PQ269271 (ITS).
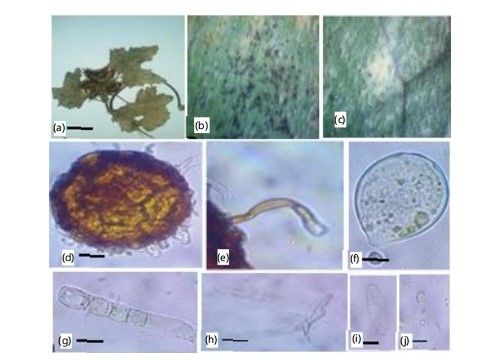
|
Mycelium amphigenous and on stems and inflorescences, in irregular patches or effuse, thin to dense, evanescent to±persistent; hyphae thin-walled, smooth or almost so, (3-)5-8(-10) μm wide; Conidiophores arising from the upper surface of hyphal mother cells, mostly towards one end of the cell but sometimes±centrally, erect, foot cells cylindrical, 30-100×10-13 μm, sometimes slightly constricted at the basal septum or slightly swollen at the very base, followed by 1-3(-4) shorter cells, forming catenescent conidia, in long chains (Fig. 10c-e); conidia ellipsoid-ovoid to doliiform, 25-45×14-22 μm (Fig. 10f-h).
The ITS sequence shows identity with the sequence of P. xanthii AB040349 from Japan. The morphological features match the description of P. xanthii given in Braun & Cook3. Senecio chrysanthemoides is being reported here as a new host record for P. xanthii from Pakistan and worldwide.
First record on Xanthium strumarium L. (Asteraceae) in Pakistan (Fig. 11a-i).
Specimen on Xanthium strumarium L. (Asteraceae) with anamorphic stage, Battagram, Hazara, Khyber Pakhtunkhwa, Pakistan 1800 m.a.s.l., 13th September, 2022, AFE094, Voucher No. HUP 16753, GenBank Accession No. PQ273711 (ITS).
Mycelium amphigenous present on inflorescences and stems, effuse or in the form of white patches, persistent, smooth hyphae, thin-walled; chasmothecia large, scattered, an irregularly polygonal shape, blackish brown in color, 77.8×74.7 μm (Fig. 11d); appendages less in number, mycelioid, long uniformly or short, septate, varies in length, 48.7×6.5 μm; pale brown in color, rough to smooth, asci subglobose to ovoid, oculus terminal, hyaline, usually immature, 55.1×45.8μm (Fig. 11e); ascospores ellipsoid-ovoid to round, late maturing, colorless when become fully mature, 12.5×11.3 μm (Fig. 11g); conidiophore appear from the top surface of mother cell, sometime centrally, infrequently in the middle, glossy, tubular, produce two-three conidia in chain, 95-143 μm long; foot cell cylindrical, 38×11-13 μm sometime slightly constricted at the basal septum followed by 1-3 shorter cell (Fig. 11h); conidia shaped in chain dolliform, 29-38×13-20 μm (Fig. 11g).
Previously, Podosphaera xanthii has been reported on Ageratum conyzoides L from Pakistan. Xanthium strumarium has been reported here as a new host record for P. xanthii from Pakistan.
In this study, new host plants for P. xanthii are being reported from different regions of Pakistan expanding the host range of this pathogen. Molecular analyses (ITS rDNA) and detailed morphological examinations of both anamorphic and teleomorphic stages provided strong evidence for the identification of P. xanthii. Large-sized ascomata observed in the Pakistani collections are consistent with the distinguishing features of P. xanthii reported in prior studies¹. The Pakistani sequences formed a sister clade with previously published reference sequences (AB046985, AB040330, PP754360, AB040293), confirming their identity3,8.
The findings align with reports by Zhang et al.28 who documented Podosphaera xanthii infecting cucurbits and other economically important crops. Recent studies by Yeh et al.8 demonstrated that environmental factors such as humidity and temperature strongly influence the pathogen’s distribution, which corresponds with our field observations in humid temperate zones of northern Pakistan. Large-sized ascomata is the distinctive character of P. xanthii that differentiates it from other members of the genus Podosphaera as reported from Korea China28,29. From this research work, it is inferred that P. xanthii not only damages economically significant plants but also poses a threat to crops, including Cucurbita. These findings emphasize the need control of disease strategies against this widely distributed pathogen in Pakistan.
CONCLUSION
This study documents the expanded host range and widespread distribution of Podosphaera xanthii across five ecological regions in Pakistan. The confirmation of its presence on ten plant species, particularly within the Asteraceae family, highlights its adaptability and potential impact on diverse plant hosts. The combined morphological and molecular analyses enhance understanding of its taxonomy and support its persistent lifecycle in varied climates. These findings provide a foundation for improved monitoring and management strategies for this economically significant pathogen.
SIGNIFICANCE STATEMENT
Powdery mildew caused by Podosphaera xanthii is a major phytopathological problem affecting a wide range of economically important crops worldwide, yet its diversity, host range, and distribution remain underexplored in many regions, including Pakistan. In this study, new host records of P. xanthii from different ecological zones of Pakistan have been reported, confirmed through detailed morphological examination and ITS based molecular analysis.
These findings significantly expand the known geographic distribution and host spectrum of P. xanthii, revealing its ecological plasticity and potential threat to both cultivated and native plant species. This study contributes valuable baseline data to the global understanding of P. xanthii diversity and host associations.
ACKNOWLEDGMENT
We pay sincere thanks to Dr. Irsa Zafar, Institute of Botany, University of the Punjab, Lahore for her valuable guidence during manuscript preparation.
REFERENCES
- Ellingham, O., G.J. Denton, J.O. Denton and R.J. Robinson, 2016. First report of Podosphaera macrospora on Heuchera in the United Kingdom. New Dis. Rep., 33: 23-23.
- Glawe, D.A., 2008. The powdery mildews: A review of the world's most familiar (yet poorly known) plant pathogens. Annu. Rev. Phytopathol., 46: 27-51.
- Braun, U. and R.T.A. Cook, 2012. Taxonomic Manual of the Erysiphales (Powdery Mildews). CBS-KNAW Fungal Biodiversity Centre, Netherlands, ISBN: 9789070351892, Pages: 707.
- Mukhtar, I., S. Mustaq and I. Khokhar, 2013. First report of powdery mildew on Dahlia (Dahlia variabilis) caused by Golovinomyces cichoracearum in Lahore, Pakistan. Australas. Plant Dis. Notes, 8: 1-3.
- Takamatsu, S., 2004. Phylogeny and evolution of the powdery mildew fungi (Erysiphales, Ascomycota) inferred from nuclear ribosomal DNA sequences. Mycoscience, 45: 147-157.
- Takamatsu, S., 2013. Molecular phylogeny reveals phenotypic evolution of powdery mildews (Erysiphales, Ascomycota). J. Gen. Plant Pathol., 79: 218-226.
- Kusch, S., M. Singh, H. Thieron, P.D. Spanu and R. Panstruga, 2023. Site-specific analysis reveals candidate cross-kingdom small RNAs, tRNA and rRNA fragments, and signs of fungal RNA phasing in the barley-powdery mildew interaction. Mol. Plant Pathol., 24: 570-587.
- Yeh, Y.W., T.Y. Wu, H.L. Wen, H.W. Jair, M.Z. Lee and R. Kirschner, 2021. Host plants of the powdery mildew fungus Podosphaera xanthii in Taiwan. Trop. Plant Pathol., 46: 44-61.
- Cunnington, J.H., S. Takamatsu, A.C. Lawrie and I.G. Pascoe, 2003. Molecular identification of anamorphic powdery mildews (Erysiphales). Australas. Plant Pathol., 32: 421-428.
- Mulpuri, S., P.K. Soni and S.K. Gonela, 2016. Morphological and molecular characterization of powdery mildew on sunflower (Helianthus annuus L.), alternate hosts and weeds commonly found in and around sunflower fields in India. Phytoparasitica, 44: 353-367.
- Kumar, S., G. Stecher and K. Tamura, 2016. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. E, 33: 1870-1874.
- Hirata, T., J.H. Cunnington, U. Paksiri, S. Limkaisang and N. Shishkoff et al., 2000. Evolutionary analysis of subsection Magnicellulatae of Podosphaera section Sphaerotheca (Erysiphales) based on the rDNA internal transcribed spacer sequences with special reference to host plants. Can. J. Bot., 78: 1521-1530.
- Cho, S.E., J.H. Park, S.H. Hong and H.D. Shin, 2013. First report of powdery mildew caused by Podosphaera xanthii on the invasive weed, Bidens pilosa, in Korea. Plant Dis., 97: 1254-1254.
- Meeboon, J. and S. Takamatsu, 2016. Notes on powdery mildews (Erysiphales) in Thailand II. Erysiphe species on Adoxaceae Anacardiaceae, Apocynaceae, Araliaceae, Aristolochiaceae, Bixaceae, Brassicaceae, Cleomaceae, Convolvulaceae, Cucurbitaceae and Euphorbiacea. Trop. Plant Pathol., 41: 357-369.
- Li, Y., Y. Gu, J. Li, M. Xu, Q. Wei and Y. Wang, 2015. Biocontrol agent Bacillus amyloliquefaciens LJ02 induces systemic resistance against cucurbits powdery mildew. Front. Microbiol., 6.
- Lee, H.B., C.J. Kim, H.Y. Mun and K.H. Lee, 2011. First report of Erysiphe quercicola causing powdery mildew on Ubame oak in Korea. Plant Dis., 95: 77-77.
- Riaz, M., N.U.S. Afshan, S. Afzal, I. Zafar, A. Saleem and A.N. Khalid, 2024. DNA barcoding and scanning electron microscopy reveals Erysiphe ahmadii sp. nov. and a new record Erysiphe populicola (Erysiphaceae, Helotiales) on salicaceous hosts from Pakistan. Microsc. Res. Tech., 87: 21-30.
- Afshan, N.S., I. Zafar and A.N. Khalid, 2021. Checklist of powdery mildew fungi (Erysiphaceae) in Pakistan. Mycotaxon, 136: 689-689.
- Afshan, N.S., J. Majeed, A.R. Niazi, S. Khanum, M. Riaz, M. Fiaz and S. Anjum, 2022. Morphological and molecular characterization of Erysiphe necator var. necator, A fungal pathogen of grapevines in Pakistan. Biologia (Lahore), 68: 18-21.
- Afshan, N.S. and M. Riaz, 2022. First record of Erysiphe aquilegiae on Clematis Montana (Himalayan Clematis) from Pakistan. Nova Hedwigia, 114: 69-76.
- Afshan, N.S., S. Afzal, M. Riaz, A. Saleem, A.R. Niazi and A.N. Khalid, 2023. Occurrence of Erysiphe friesii var. dahurica on purple buckthorn (Rhamnus purpurea) from Pakistan. Can. J. Plant. Pathol., 45: 427-432.
- Afzal, S., N.U.S. Afshan, S. Seerat, I. Zafar, M. Jabeen, M. Riaz and A.N. Khalid, 2022. New records of Podosphaera species (Ascomycota: Erysiphaceae) on members of Rosaceae from Pakistan. Nova Hedwigia, 115: 417-429.
- Darsaraei, H., N.U.S. Afshan, S. Afzal, I. Zafar and A. Anwar et al., 2023. A re-assessment of Erysiphe polygoni: Phylogenetic analyses suggest an undescribed species. Sydowia, 75: 233-242.
- Zafar, I., N.S. Afshan, J. Majeed, A.R. Niazi and A.N. Khalid, 2022. Erysiphe bistortae sp. nov. on Bistorta amplexicaulis from Pakistan. Mycotaxon, 137: 883-891.
- Zafar, I., S. Afzal, N.U.S. Afshan and A.N. Khalid, 2023. Golovinomyces artemisiae and Golovinomyces sonchicola, additions to the Erysiphales of Pakistan. Biol. Bull., 50: 348-355.
- Zafar, I., N.U.S. Afshan, A.N. Khalid and M. Götz, 2023. Erysiphe machilicola: A new powdery mildew species on Duthie’s bay tree. Phytotaxa, 594: 283-291.
- Jabeen, M., N.U.S. Afshan, S. Afzal, A.N. Khalid and A. Saleem, 2025. Morphological and molecular characterization of Podosphaera hyalina comb. nov. parasitizing creeping sibbaldia (Sibbaldia procumbens) in Pakistan. Can. J. Plant. Pathol.
- Zhang, S., J. Liu, B. Xu and J. Zhou, 2021. Differential responses of Cucurbita pepo to Podosphaera xanthii reveal the mechanism of powdery mildew disease resistance in pumpkin. Front. Plant Sci., 12.
- Choi, I.Y., J.H. Kim, M.J. Uhm, S.E. Cho and H.D. Shin, 2018. First report of powdery mildew caused by Podosphaera xanthii on okra in Korea. Plant Dis., 102: 1663-1663.
How to Cite this paper?
APA-7 Style
Afshan,
N., Jabeen,
M., Akbar,
A., Khalid,
M., Saleem,
A., Altaf,
R., Afzal,
S., Fiaz,
M., Khalid,
A.N. (2025). Geographic Distribution and Genetic Diversity of Podosphaera xanthii in Pakistan. Asian Journal of Plant Pathology, 19(1), 81-95. https://doi.org/10.3923/ajpp.2025.81.95
ACS Style
Afshan,
N.; Jabeen,
M.; Akbar,
A.; Khalid,
M.; Saleem,
A.; Altaf,
R.; Afzal,
S.; Fiaz,
M.; Khalid,
A.N. Geographic Distribution and Genetic Diversity of Podosphaera xanthii in Pakistan. Asian J. Plant Pathol. 2025, 19, 81-95. https://doi.org/10.3923/ajpp.2025.81.95
AMA Style
Afshan
N, Jabeen
M, Akbar
A, Khalid
M, Saleem
A, Altaf
R, Afzal
S, Fiaz
M, Khalid
AN. Geographic Distribution and Genetic Diversity of Podosphaera xanthii in Pakistan. Asian Journal of Plant Pathology. 2025; 19(1): 81-95. https://doi.org/10.3923/ajpp.2025.81.95
Chicago/Turabian Style
Afshan, Najam-ul-Sehar, Muhammada Jabeen, Afshan Akbar, Muhammadah Khalid, Arooma Saleem, Rabia Altaf, Saliha Afzal, Muhammad Fiaz, and Abdul Nasir Khalid.
2025. "Geographic Distribution and Genetic Diversity of Podosphaera xanthii in Pakistan" Asian Journal of Plant Pathology 19, no. 1: 81-95. https://doi.org/10.3923/ajpp.2025.81.95

This work is licensed under a Creative Commons Attribution 4.0 International License.